Реверсия заряда: Потенциал действия. Ионный механизм возникновения потенциала действия
Потенциал действия. Ионный механизм возникновения потенциала действия
⇐ ПредыдущаяСтр 2 из 19Следующая ⇒
Потенциалом действия (ПД)называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения.Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды.
В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе.Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы.
| В ПД выделяют следующие фазы (рис. 3):
■ деполяризации; ■ реполяризации. |
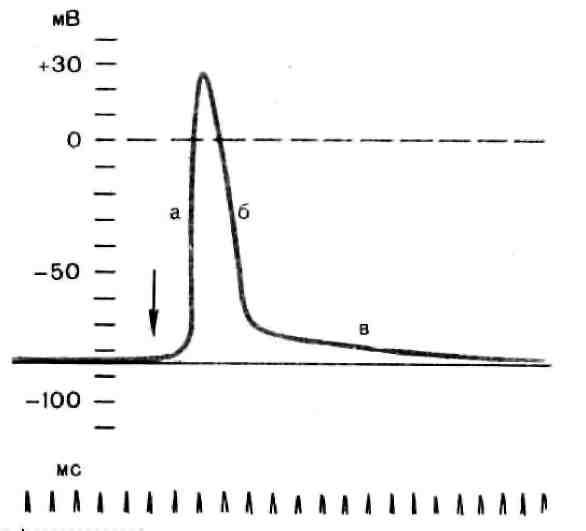
Рис. 3. Потенциал действия скелетного мышечного волокна,зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации, б – фаза реполяризации, в – фаза следовой деполяризации (отрицательный следовой потенциал). Момент нанесения раздражения показан стрелкой.
Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала.Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 – +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации.
Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами.Различают два вида следовых потенциалов – следовую деполяризацию и следовую гиперполяризацию.
Ионный механизм возникновения ПД.Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К+ из цитоплазмы во внешний раствор превышает противоположно направленный поток Na+. Поэтому наружная сторона мембраны в покое имеет положительный потенциал по отношению к внутренней.
При действии на клетку раздражителя проницаемость мембраны для Na+ резко повышается и становится примерно в 20 раз больше проницаемости для K+. Поэтому поток Na+ из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутренняя сторона мембраны в месте возбуждения становится заряженной положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе ПД (фазе деполяризации).
Повышение проницаемости мембраны для Na+ продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na+ вновь понижается, а для K+ возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией.В результате инактивации поток Na+ внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K+ из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД.
ПД возникает при деполяризации поверхностной мембраны.Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению ионов Na внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные изменения на мембране (локальный ответ).При увеличении силы раздражения, когда достигнут порог возбудимости, изменения мембранного потенциала достигают критического уровня деполяризации (КУД). Например, величина потенциала покоя равна -70 мВ, КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т.е. на -20 мВ снизить ее исходный потенциал покоя. Только при достижении КУД наблюдается резкое изменение мембранного потенциала, которое регистрируется в виде ПД. Таким образом, основное условие возникновения потенциала действия – это снижение мембранного потенциала до критического уровня деполяризации.
В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами:
■ избирательностью (селективностью) по отношению к определенным ионам;
■ электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала.
Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны.
Активный и пассивный ионный транспорт.В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na+ и K+ по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии.
Таким образом, в живой клетке существует две системы движения ионов через мембрану.
Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны.
Второй осуществляется против концентрационного градиента. Он состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом.Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт — результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения.
Проведение возбуждения
Нервный импульс (потенциал действия) обладает способностью распространяться вдоль по нервным и мышечным волокнам.
В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5 – 6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения.
Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны – положительный) и соседним невозбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи – так называемые местные токи.В результате развивается деполяризация соседнего участка, увеличение его ионной проницаемости и появление потенциала действия. В исходной же зоне возбуждения восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т.д. Таким образом, с помощью местных токов происходит распространение возбуждения на соседние участки нервного волокна, т.е. проведение нервного импульса.По мере проведения амплитуда потенциала действия не уменьшается,т.е. возбуждение не затухает даже при большой длине нерва.
В процессе эволюции с переходом от безмякотных нервных волокон к мякотным (покрытым миелиновой оболочкой) произошло существенное повышение скорости проведения нервного импульса. Для безмякотных волокон характерно непрерывное проведение возбуждения, которое охватывает последовательно каждый соседний участок нерва. Мякотные же нервы почти полностью покрыты изолирующей миелиновой оболочкой. Ионные токи в них могут проходить только в оголенных участках мембраны -перехватах Ранвье, лишенных этой оболочки. При проведении нервного импульса потенциал действия перескакивает от одного перехвата к другому и может охватывать даже несколько перехватов. Такое проведение поучило название сальтоторного (лат. сальто – прыжок). При этом повышается не только скорость, но и экономичность проведения. Возбуждение захватывает не всю поверхность мембраны волокна, а лишь небольшую ее часть. Следовательно, меньше энергии тратится на активный транспорт ионов через мембрану при возбуждении и в процессе восстановления.
Скорость проведения в разных волокнах различна. Более толстые нервные волокна проводят возбуждение с большей скоростью: у них расстояния между перехватами Ранвье больше и длиннее скачки. Наибольшую скорость проведения имеют двигательные и проприоцептивные афферентные нервные волокна — до 100 м/с. В тонких симпатических нерв ных волокнах (особенно в немиелинизированных волокнах) скорость проведения мала — порядка 0,5 — 15 м/с.
Во время развития потенциала действия мембрана полностью теряет возбудимость. Это состояние называют полной невозбудимостью, или абсолютной рефрактерностью.За ним следует относительная рефрактерность, когда потенциал действия может возникать лишь при очень сильном раздражении. Постепенно возбудимость восстанавливается до исходного уровня.
Законы проведения возбуждения в нервах:
1. Проведение импульсов возможно лишь при условии анатомической и физиологической целостности волокна.
2. Двустороннее проведение: при раздражении нервного волокна возбуждение распространяется по нему и в центробежном, и в центростремительном направлениях.
3. Изолированное проведение: в периферическом нерве импульсы распространяются по каждому волокну изолированно, т.е. не переходя с одного волокна на другое и оказывая действие только на те клетки, с которыми контактируют окончания данного нервного волокна.
13. Дайте определение гомеостаза.
14. Назовите основные пути регуляции различных функций у высокоорганизованных животных и человека.
15. Кем и когда было открыто «животное электричество»?
16. Какие ткани относятся к возбудимым? Почему они так называются?
17. Назовите основные функциональные характеристики возбудимых тканей.
18. Что называют порогом возбудимости?
19. От каких факторов зависит величина порога?
20. Что такое лабильность? Кем было выдвинуто понятие лабильности, какие свойства возбудимых тканей оно характеризует?
21. Что называют мембранным потенциалом (потенциалом покоя)?
22. Чем обусловлено наличие электрических потенциалов в живых клетках?
23. В каких случаях говорят о деполяризации (или гиперполяризации) клеточной мембраны?
24. Какую роль в формировании потенциала покоя играет калий-натриевый насос мембраны?
25. Что называют потенциалом действия? Какова его роль в нервной системе?
26. Что лежит в основе возникновения потенциала действия?
27. Охарактеризуйте фазы потенциала действия.
28. Что называют реверсией мембранного потенциала?
29. Опишите ионный механизм возникновения потенциала действия.
30. Что понимают под натриевой инактивацией?
31. Что такое критический уровень деполяризации?
32. Какими свойствами обладают ионные каналы клеточной мембраны?
33. Охарактеризуйте два типа ионного транспорта в клетке:
■ пассивный;
■ активный.
Модуль 1 ОБЩАЯ ФИЗИОЛОГИЯ ЦНС
Реверсивная зарядка смартфонов — что это и как работает?
Как новые технологии позволяю зарядить один смартфон от другого без проводов и розетки?
В последние 3-4 года новым трендом среди производителей смартфонов стала беспроводная зарядка. Первыми, кто реализовали эту технологию, стали разработчики Android-флагманов. В 2017 году в гонку вступила компания Apple, интегрировав эту опцию в iPhone 8 и iPhone X. Сейчас на флагманских телефонах используют новую технологию, которая называется реверсивная зарядка.
Что такое реверсивная зарядка?
Первый смартфон, который начал использовать эту технологию — Huawei Mate 20 Pro. Дальше активным развитием этой идеи занялся один из самых крупных игроков на рынке мобильных устройств — Samsung. Южнокорейская компания назвала эту функцию PowerShare и внедрила ее в Note10, Note10+ и линейку смартфонов Galaxy S10.
Владельцы этих гаджетов могут заряжать один телефон от другого без помощи зарядного устройства и проводов. Причем это может быть не только смартфоны — например, от телефона можно зарядить умные часы или беспроводные наушники. Таким образом, смартфон превращается в автономную зарядную станцию. К задней крышке гаджета можно приложить другой телефон или умные аксессуары (тоже задней крышкой), после чего начнется процесс зарядки.

Чтобы реализовать эту технологию, производителям приходится увеличивать объем аккумулятора. Реверсивная передача питания дает возможность передавать часть заряда батареи на разрядившееся устройство.
Чтобы воспользоваться этой технологией, нужно активировать опцию на смартфоне через настройки. Далее гаджеты необходимо соединить друг с другом задней панелью. Если телефон в чехле, его нужно снять. Оба мобильных устройства должны быть включены. Нельзя помещать металлические предметы между двумя гаджетами. Также не рекомендуется класть телефоны на металлическую поверхность — из-за этого могут возникнуть сбои.
Если уровень заряда батареи телефона ниже установленного, опция будет отключена в автоматическом режиме. Главный недостаток этого метода зарядки — он очень медленный. Также нельзя зарядить смартфон на 100%, так как одно устройство передает собственную энергию.
В определенный момент возможность реверсивной зарядки отключится — когда у телефона, который используется в качестве автономной зарядной станции, закончится собственный заряд батареи. Как правило, передача питания отключается, когда заряда аккумулятора становится меньше 30%. Таким образом производители решают проблему полного разряда аккумулятора во время зарядки другого гаджета. У Huawei функция отключается, когда передающий заряд смартфон заряжен менее чем на 20%.

В Galaxy S10 инженеры реализовали функцию двойной передачи питания. Гаджет можно подключить к проводной зарядке и активировать функцию PowerShare. После этого можно использовать устройство как полноценное автономное ЗУ, например, положить телефон на стол и заряжать умные часы или другой полезный мобильный аксессуар, который поддерживает технологию беспроводной зарядкой.
Чтобы отключить зарядку, нужно просто «отделить» устройства друг от друга. Если смартфон не может найти подходящий гаджет, функция реверсивной зарядки отключится автоматически — через 30-40 секунд.
 Загрузка…
Загрузка…
11. Изменение проницаемости мембраны при развитии волны возбуждения.
Мембранный
потенциал. Снаружи мембрана заряжена
положительно, а изнутри — отрицательной.
70-80 мВ.
Ток
повреждения
— это разность заряда между неповрежденным
и поврежденным. Поврежденный заряжен
отрицательно, относительно целой.
Метаболический
ток
— это разность понетциалов вследствии
неодинаковой интенсивности обменных
процессов.
Происхождение
мембранного потенциала объясняют с
точки зрения мембранно-ионной
теории,
которая учитывает неодинаковую
проницаемость мембраны для ионов и
разный состав ионов во внутриклеточной
и межклеточной жидкости. Установлено,
что и внутриклеточная и межклеточная
жидкость имеют одинаковое количество
и положительных и отрицательных ионов,
однако состав разный. Внешняя жидкость:
Na+, Cl-Внутренняя жидкость: K+, A-(органические
анионы)
В
состоянии покоя мембрана по разному
проницаема для ионов.
Наибольшая проницаемость у калия, затем
идет натрий и хлор. Для органических
анионов мембраны не проницаемы.
Вследствие
повышенной проницаемости для ионов
калия, они выходят из клетки. В результате
чего внутри скапливаются орг. анионы.
В результате создается разница потенциалов
(диффузионный калиевый потенциал),
который идет до тех пор, пока он может
выходить.
Расчетный
калиевый потенциал равен -90 мВ. А
практический потенциал равен -70 мВ. Это
говорит о том, что в создании потенциала
участвует и другой ион.
Для
того, чтобы сдерживать потенциал в
мембране, клетка должна работать, ибо
перемещение ионов калия из клетки, а
натрия в клетку, привело бы к нарушению
равенства знака. Мембраны поляризованы.
Снаружи заряд будет положительным, а
снаружи — отрицательным.
Состояние
электрического заряда мембраны.
Реверсия
или овершут
— изменение знака заряда. Возвращение
к исходному заряду — реполяризация.
Токи
при возбуждении.
При
действии раздражителя на мембрану
происходит кратковременное возбуждение.
Процесс возбуждения является локальным
и распространяется вдоль мембраны, а
потом деполяризуется. По мере движения
возбуждения деполяризуется новый
участок мембраны и т.д. Ток действия
является двухфазным током.
12.Законы ритмического раздражения — оптимум и пессимум. Парабиоз и его стадии.
Повышение
частоты и силы раздражения до известного
предела вызывает увеличение высоты
тетанического сокращения скелетной
мышцы. Наиболее благоприятная частота
нервных импульсов, поступающих в
скелетную мышцу, вызывает наибольшую
высоту тетануса. Эта частота называется
оптимальной, или оптимумом
частоты.
Оптимуму частоты соответствует такая
частота, при которой каждое последующее
раздражение застает скелетную мышцу в
состоянии наибольшей возбудимости,
наблюдающейся в экзальтационной фазе.
Поэтому высота каждого одиночного
сокращения возрастает. Наоборот, если
каждое последующее раздражение застает
скелетную мышцу в фазе абсолютной
рефрактерности, то тетаническое
сокращение мышцы резко уменьшается или
не наступает. Эта чрезмерно большая
частота — наихудшая, пессимальная, или
пессимум
частоты.
Каждая волна возбуждения не только
вызывает сокращение скелетной мышцы,
но и сопровождается изменениями ее
возбудимости и лабильности. Поэтому
последующая волна возбуждения застает
скелетную мышцу либо в состоянии
экзальтационной фазы, обусловленной
предыдущим раздражением (оптимум
частоты), либо в абсолютной рефрактерной
фазе, или интервале невозбудимое™,
созданном предыдущим раздражением
(пессимум частоты). Оптимум частоты
соответствует высокому уровню лабильности
нерва и мышцы, а пессимум частоты —
низкому уровню лабильности нерва, даже
более низкому, чем лабильность мышцы.
В результате предыдущих раздражений
при пессимуме частоты лабильность
нервно-мышечного препарата резко
снижается и полностью задерживается
переход волн возбуждения с нерва на
мышцу, наступает торможение, тетанус
отсутствует. Наиболее благоприятная
сила раздражения, вызывающая максимальное
тетаническое сокращение скелетной
мышцы, называется оптимумом силы.
Уменьшение силы раздражения снова
вызывает тетаническое сокращение
скелетной мышцы. Оптимум
и пессимум силы
— также результат изменений возбудимости
и лабильности, вызываемых предыдущими
раздражениями.
Парабиоз-функциональные
изменения, наступающие в нерве в ответ
на действие сильных и длительно
действующих альтерирующих раздражителей.
Открытие П. и сам термин принадлежат Н.
Е. Введенскому.
Механизм генерации потенциала действия (ПД) — Студопедия
Дальнейшие исследования Ходжкина и Хаксли показали, что при возбуждении аксона кальмара возникает быстрое колебание мембранного потенциала, которое на экране осциллографа имело форму пика (spike). Они назвали это колебание потенциалом действия (ПД). Так как электрический ток для возбудимых мембран является адекватным раздражителем, ПД можно вызвать, поместив на наружную поверхность мембраны отрицательный электрод – катод, а внутреннюю положительный — анод. Это приведет к снижению величины заряда мембраны – ее деполяризации. При действии слабого допорогового тока происходит пассивная деполяризация, т.е. возникает катэлектротон. Если силу тока увеличить до определенного предела, то в конце периода его воздействия на плато катэлектротона появится небольшой самопроизвольный подъём – местный или локальный ответ. Он является следствием открывания небольшой части натриевых каналов, находящихся под катодом. При токе пороговой силы МП снижается до критического уровня деполяризации (КУД), при котором начинается генерация потенциала действия. Он находится для нейронов примерно на уровне – 50 мВ.
На кривой потенциала действия выделяют следующие фазы:
1. Локальный ответ (местная деполяризация), предшествующий развитию ПД.
2. Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше нуля. Поэтому мембрана приобретает противоположный заряд – внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мс.
3. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мс.
4. Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мс.
5. Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мс.
Амплитуда ПД скелетных мышц в среднем 120-130 мВ, нейронов 80-90 мВ, гладкомышечных клеток 40-50 мВ. При возбуждении нейронов ПД возникает в начальном сегменте аксона – аксонном холмике.
Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые, и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных ворот зависит величина КУД (чем она выше, тем ниже КУД, и наоборот).
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию. Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной после генерации ПД калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия.
Изменяя проводимость быстрых натриевых и калиевых каналов, можно влиять на генерацию ПД, а, следовательно, на возбуждение клеток. При полной блокаде натриевых каналов, например, ядом рыбы тетродонта – тетродотоксином, клетка становится невозбудимой. Это используется в клинике. Такие местные анестетики, как новокаин, дикаин, лидокаин тормозят переход натриевых каналов нервных волокон в открытое состояние. Поэтому проведение нервных импульсов по чувствительным нервам прекращается, наступает обезболивание (анестезия) органа. При блокаде калиевых каналов затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, т.е. восстановление МП. Поэтому удлиняется фаза реполяризации. Этот эффект блокаторов калиевых каналов также используется в клинической практике. Например, один из них хинидин, удлиняя фазу реполяризации кардиомиоцитов, урежает сердечные сокращения и нормализует сердечный ритм.
Также следует отметить, что чем выше скорость распространения ПД по мембране клетки или ткани, тем выше ее проводимость.
2_1 Мембранный потенциал покоя кратко (доцент Сазонов В.Ф.)
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
— На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
— На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка — нейрон.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом перемещаются электрические заряды на нервных клетках к как за счёт этого на мембране появляется электрический потенцила. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов — формирование потенциала покоя.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя — это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
«Потенциал» — это возможность, он сродни понятию «потенция». Электрический потенциал мембраны — это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы — ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные — положительно (+).
Таким образом, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках — это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать «плюсики», т.е. положительно заряженные ионы, а внутри — «минусики», т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки — отрицательные «персонажи». ((
Сущность
Сущность потенциала покоя — это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки — «отрицательность», а снаружи — «положительность».
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны , (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях — и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
1) Через ионные насосы-обменники натрий насильно выводится из клетки, а калий затаскивается в клетку.
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне — 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Попробую рассказать коротко, откуда берётся мембранный потенциал покоя в нервных клетках — нейронах. Ведь, как всем теперь известно, наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц — анионов и недостаток положительных частиц — катионов.
И вот тут исследователя и студента поджидает одна из логических ловушек: внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот — из-за потери некоторого количества положительных частиц (катионов).
И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов — катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомню, что это ионы натрия — Na+ и калия — K+.
Натрий-калиевый насос
А всё дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют «собственный» натрий клетки на наружный «чужой» калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Чтобы легче было запомнить, образно можно сказать так: «Клетка любит калий!» (Хотя об истинной любви здесь не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его и так полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от -10 до -70 mV, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки . А в экспериментах на мультипотентных мезенхимальных стромальных клетках (ММСК) костного мозга человека искусственная деполяризация ингибировала дифференцировку клеток (Fischer-Lougheed J., Liu J.H., Espinos E. et al. Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology 2001; 153: 677-85; Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology 1998; 510: 467-76; Sundelacruz S., Levin M., Kaplan D.L. Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. Plos One 2008; 3).
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка «заряжается любовью».
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Как ни странно, но результат этих двух видов любви — пустота!
Именно она, пустота, создаёт в клетке отрицательный электрический заряд — потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал — возможность затягивать внутрь клетки положительно заряженные частицы (ионы).
2. Ионный натриевый потенциал — возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какие-нибудь другие).
3. Ионный калиевый потенциал — возможновть выталкивать из клетки ионы калия (и именно калия, а не какие-нибудь другие).
1. Дефицит натрия (Na+) в клетке.
2. Избыток калия (K+) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь «полезет» этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выравнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно — концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
От химического — к электрическому
А теперь — самое главное, следите за излагаемой мыслью! Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и «плюсики» (положительные заряды). На их месте в клетке остаются «минусы» (отрицательные заряды). Это и есть мембранный потенциал покоя!
Мембранный потенциал покоя — это дефицит положительных зарядов внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Заключение

Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Потенциал покоя — отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть — это примерно -10 милливольт, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает «плюсиков» с натрием, чем закачивает обратно с калием).
2. Вторая часть — это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до -70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в -90 милливольт. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне -70 милливольт.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки «лишнего» калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть «мембранный потенциал покоя». Он измеряется внутри клетки и составляет обычно -70 мВ.
Выводы
Говоря образно, «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками».
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше «плюсиков» (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки — неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Итак:
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это — разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Видео: Мембранный потенциал покоя (Resting membrane potential)
© 2009-2018 Сазонов В.Ф. © 2009-2016 kineziolog.bodhy.ru, © 2016-2018 kineziolog.su,
Потенциал действия | Info-Farm.RU
Потенциал действия (ПД) — это кратковременные амплитудные изменения мембранного потенциала покоя (МПС), возникающие при возбуждении живой клетки. По сути это электрический разряд — быстрая кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона или мышечного волокна), в результате которого внешняя поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, который играет сигнальную (регуляторную) роль.
Общая характеристика
Потенциалы действия могут отличаться по своим параметрам в зависимости от типа клетки и даже на разных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Все же, в основе любого потенциала действия лежат следующие явления:
- «Мембрана живой клетки поляризована» — ее внутренняя поверхность заряжена отрицательно по отношению к наружной благодаря тому, что в растворе у ее внешней поверхности находится большее количество положительно заряженных частиц (катионов), а у внутренней поверхности — большее количество отрицательно заряженных частиц (анионов).
- «Мембрана имеет избирательную проницаемость ‘- ее проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
- «Мембрана возбудимой клетки способна быстро менять свою проницаемость ‘для определенного вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю
Первые два свойства характерны для всех живых клеток. Третья же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Основной математической моделью, описывающей генерацию и передачу потенциала действия, является модель Ходжкина-Хаксли.
Фазы
Можно четко выделить пять фаз развития ПД:
Нарастание (деполяризация)
Возникновение потенциала действия (ПД) связано с увеличением проницаемости мембраны для ионов натрия (в 20 раз по сравнению с проницаемостью для К +, и в 500 раз по сравнению с исходной проницаемостью Na +) и последующим усилением диффузии этих ионов по концентрационном градиенту внутрь клетки, приводит к изменению (уменьшение) мембранного потенциала. Уменьшение мембранного потенциала приводит к увеличению проницаемости мембраны для натрия путем открытия потенциал-зависимых натриевых каналов, а увеличение проницаемости сопровождается усилением диффузии натрия в цитоплазму, что вызывает еще более значительную деполяризацию мембраны. Благодаря наличию положительной обратной связи деполяризация мембраны при возбуждении происходит с ускорением и поток ионов натрия в клетку все время растет. Интенсивность же потока ионов калия, направленного из клетки наружу, в первые моменты возбуждения остается в начале. Усиленный поток положительно заряженных ионов натрия внутрь клетки вызывает сначала исчезновение избыточного отрицательного заряда на внутренней поверхности мембраны, а затем приводит к перезарядки мембраны. Поступления ионов натрия происходит до тех пор, пока внутренняя поверхность мембраны не приобретет положительный заряд, достаточный для уравновешивания градиента концентрации натрия и прекращение его дальнейшего перехода внутрь клетки. Натриевый возникновения ПД подтверждают опыты с изменением внешней и внутренней концентрации этого иона. Было показано, что десятикратном изменении концентрации ионов натрия во внешнем или внутреннем среде клетки, соответствует изменение ПД на 58 мВ. При полном удалении ионов натрия из окружающей клетку жидкости ПД ни возникал. Таким образом, установлено, что ПД возникает в результате избыточной, по сравнению с покоем, диффузии ионов натрия из окружающей жидкости внутрь клетки. Период, в течение которого проницаемость мембраны для ионов натрия при открытии натриевых каналов растет, является небольшим (0,5-1 мс) вслед за этим наблюдается повышение проницаемости мембраны для ионов калия благодаря открытию потенциал-зависимых калиевых каналов, и, следовательно, усиление диффузии этих ионов из клетки наружу.
- Принцип «все или ничего»
- Согласно закону «все-или-ничего» мембрана клетки возбудимой ткани или не отвечает стимул совсем, или отвечает с максимально возможной для нее на данный момент силой. Действие раздражителя обычно приводит к локальной деполяризации мембраны. Это вызывает открытие натриевых каналов, которые чувствительны к изменениям потенциала, а через это — увеличивает натриевую проводимость, что приводит к еще большей деполяризации. Существование такой обратной связи обеспечивает регенеративную (возобновляемую) деполяризацию клеточной мембраны. Величина потенциала действия зависит от силы раздражителя, а сам он возникает только в том случае, когда деполяризация превышает некоторый определенный для каждой клетки предельный уровень. Это явление получило название «все или ничего». Однако, если деполяризация составляет 50-75% от предельной величины, то в клетке может возникнуть локальный ответ, амплитуда которой значительно ниже амплитуду потенциала действия. Отсутствие потенциала действия при пидграничному уровне деполяризации объясняется тем, что при этом недостаточно увеличивается натриевая проницаемость, чтобы вызвать регенеративную деполяризацию. Уровень деполяризации, который возникает при этом, не вызывает открытие новых натриевых каналов, поэтому натриевая проводимость быстро уменьшается, и в клетке снова устанавливается потенциал покоя.
Овершут
Деполяризация мембраны приводит к реверсии мембранного потенциала (МП становится положительным). В фазу овершута Na + -ток начинает стремительно снижаться, что связано с инактивацией потенциал-зависимых Na + -каналов (время открытого состояния — судьбы миллисекунды) и исчезновением электрохимического градиента Na +.
- Рефрактерность
- Одним из последствий исчезновения градиента Na + является рефрактерность мембраны — временная неспособность отвечать на раздражитель. Если раздражитель возникает сразу после прохождения потенциала действия, то возбудимость не возникнет ни при силе раздражителя на уровне порога, ни при значительно более сильное раздражителю. Такое положение полной невозбудимости называется абсолютным рефрактерным периодом. За ним следует относительный рефрактерный период, когда надпороговый раздражитель может вызвать потенциал действия со значительно меньшей амплитудой чем в норме. Потенциал действия привычной амплитуды при действии порогового раздражителя можно вызвать только после нескольких миллисекунд после предварительного потенциала действия. Абсолютный рефрактерный период ограничивает максимальную частоту генерации потенциалов действия.
Реполяризация
Увеличение ионного потока калия, направленного из клетки наружу, приводит к уменьшению мембранного потенциала, в свою очередь обусловливает уменьшение проницаемости мембраны для ионов натрия, что, как указывалось, является функцией мембранного потенциала. Таким образом, второй этап характеризуется тем, что поток ионов калия из клетки наружу растет, а встречный поток ионов натрия уменьшается. Такая реполяризация мембраны продолжается, пока не произойдет восстановление потенциала покоя — реполяризация мембраны. После этого проницаемость для ионов калия также падает до исходной величины. Внешняя поверхность мембраны за счет положительно заряженных ионов калия, вышедших в среду, вновь приобретает положительный потенциала относительно внутреннего.
Следовая деполяризация и гиперполяризация
В конечной фазе происходит замедление восстановления мембранного потенциала покоя, и при этом регистрируются следовые реакции в виде следовой деполяризации и гиперполяризации, обусловлены медленным восстановлением исходной проницаемости для ионов К +.
Распространение
Распространение в немиелинизированные волокне
В немиелинизированные (без`мякотному) нервном волокне ПД распространяется от точки к точке, поскольку возбуждение можно зарегистрировать как такое, что постепенно «бежит» по всему волокну от места своего возникновения. Ионы натрия, входящих внутрь возбуждении участка, служат источником электрического тока для возникновения ПД в прилегающих участках. В этом случае импульс возникает между деполяризована участком мембраны и ее невозбужденном участком. Разность потенциалов здесь во много раз выше, чем необходимо для того, чтобы деполяризация мембраны достигла предельного уровня. Скорость распространения импульса в таких волокнах 0,5-2 м / с
Распространение в миелинизированные волокне
Нервные отростки большинства соматических нервов миелинизированные. Только очень незначительные их участки, так называемые перехвата узла (перехват Ранвье), покрытые обычной клеточной мембраной. Такие нервные волокна характеризуются тем, что на мембране только в перехватах размещении потенциал-зависимые ионные каналы. Кроме того, эта оболочка повышает электрическое сопротивление мембраны. Поэтому при сдвиге мембранного потенциала ток проходит через мембрану перехватывающих участка, то есть прыжками (сальтаторно) от одного перехвата к другому, что позволяет увеличить скорость проведения нервного импульса, которая составляет от 5 до 120 м / с. Причем потенциал действия, который возник в одном из перехватов Ранвье, вызывает потенциалы действия в соседних перехвата за счет возникновения электрического поля, которое вызывает начальную деполяризацию в этих перехватов. Параметры ЭДС поля и дистанция его эффективного действия зависят от кабельных свойств аксона.
| Тип | Диаметр (мкм) | Миелинизация | Скорость проведения (м / с) | Функциональное назначение |
|---|---|---|---|---|
| А alpha | 12-20 | сильная | 70-120 | Подвижные волокна соматической НС; чувствительные волокна проприорецепторов |
| А beta | 5-12 | сильная | 30-70 | Чувствительные волокна рецепторов кожи |
| А gamma | 3-16 | сильная | 15-30 | Чувствительные волокна проприорецепторов |
| А delta | 2-5 | сильная | 12-30 | Чувствительные волокна терморецепторов, ноцицепторов |
| В | 1-3 | слабая | 3-15 | Преганглионарные волокна симпатической НС |
| С | 0,3-1,3 | отсутствует | 0,5-2,3 | Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов некоторых механорецепторов |
Распространение потенциала действия между клетками
В химическом синапсе после того, как волна потенциала действия доходит нервного окончания, она вызывает высвобождение нейротрансмиттеров из пресинаптических пузырьков в синаптическую щель. Молекулы медиатора, высвобождаемых с пресинапса, связываются с рецепторами на постсинаптической мембране, в результате чего в рецепторных макромолекулах открываются ионные каналы. Ионы, начинают поступать внутрь постсинаптической клетки через открытые каналы, изменяют заряд ее мембраны, что приводит к частичной деполяризации мембраны и, как следствие, провоцирование генерации постсинаптической клетки потенциала действия.
В электрическом синапсе отсутствует «посредник» передачи в виде нейромедиатора. Зато клетки соединены между собой с помощью специфических протеиновых тоннелей — конексонив, поэтому ионные токи, с пресинаптической клетки могут стимулировать постсинаптическую клетку, вызывая зарождения в ней потенциала действия. Благодаря такому строению, потенциал действия может распространяться в обе стороны и значительно быстрее, чем через химический синапс.
Схема процесса передачи нервного сигнала в химическом синапсе
Схема строения электрического синапса
Потенциал действия в различных типах клеток
Потенциал действия в мышечных тканях
Потенциал действия в скелетных мышечных клетках аналогичный потенциала действия в нейронах. Потенциал покоя в них как правило -90мВ, что меньше, чем потенциал покоя типовых нейронов. Потенциал действия мышечных клеток длится примерно 2-4 мс, абсолютный рефрактерный период составляет примерно 1-3 мс, а скорость проводимости вдоль мышц примерно 5 м / с.
Потенциал действия в сердечных тканях
Потенциал действия клеток рабочего миокарда состоит из фазы быстрого деполяризации, начальной быстрой реполяризации, которая переходит в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации. Фаза быстрой деполяризации обусловлена резким повышением проницаемости мембраны для ионов натрия, вызывает быстрый входящий натриевый ток, при достижении мембранного потенциала 30-40 мВ инактивируется и в дальнейшем главную роль играют кальциевый ионный ток. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный Деполяризующий входящий кальциевый ток.
Потенциал действия в сердечной ткани играет важную роль в координации сокращений сердца.
Молекулярные механизмы возникновения потенциала действия
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основанные главным образом на поведении потенциал-зависимых натриевых (Na +) и калиевых (K +) каналов. Начальная фаза ПД формируется входным натриевым током, позже открываются калиевые каналы и выходной K + -ток возвращает потенциал мембраны к исходному уровню. Начальное концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: в Na + -каналов основных состояния трех — закрытый, открытый и инактивированный (в реальности все сложнее, но этих трех состояний достаточно для описания), в K + каналов два — закрытый и открытый.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и рассчитывается через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Алан Ллойд Ходжкин и Эндрю Хаксли.
- Проводимость для калия G K на единицу площади [S / cm²]
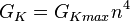 , , 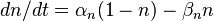 |
| где: |
| an — Коэффициент трансфера из закрытого в открытое состояние для K + каналов [1 / s]; |
| bn — Коэффициент трансфера из открытого в закрытое состояние для K + каналов [1 / s]; |
| n — Фракция К + каналов в открытом состоянии; |
| (1 — n) — Фракция К + каналов в закрытом состоянии |
- Проводимость для натрия G Na на единицу площади [S / cm²]
рассчитать сложнее, поскольку, как уже было упомянуто, в потенциал-зависимых Na + каналов, кроме закрытого / открытого состояний, переход между которыми параметром, еще инактивированный / никак инактивированный состояния, переход между которыми описывается через параметр
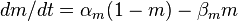 , , | 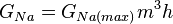 , , |
| где: | 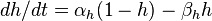 где: где: |
| am — Коэффициент трансфера из закрытого в открытое состояние для Na + каналов [1 / s]; | ah — Коэффициент трансфера из инактивированного в не-инактивированный состояние для Na + каналов [1 / s]; |
| bm — Коэффициент трансфера из открытого в закрытое состояние для Na + каналов [1 / s]; | bh — Коэффициент трансфера из не-инактивированного в инактивированный состояние для Na + каналов [1 / s]; |
| m — Фракция Na + каналов в открытом состоянии; | h — Фракция Na + каналов в не-инактивированном состоянии; |
| (1 — m) — Фракция Na + каналов в закрытом состоянии | (1 — h) — Фракция Na + каналов в инактивированном состоянии. |
Методы исследования
История
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю.Бернштейном
В 1902 году Юлиус Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К +, и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазме мышцы и окружающей средой, который составил около — 70 мВ. Согласно теории Ю.Бернштейна, при возбуждении клетки ее мембрана повреждается, и ионы К + выходят из клетки по концентрационном градиента до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя.
Эту модель развили в своей работе 1952 Алан Ллойд Ходжкин и Эндрю Хаксли в которой описали электрические механизмы, обусловливающие генерацию и передачу нервного сигнала в гигантском аксоне кальмара. За это авторы модели получили Нобелевскую премию в области физиологии и медицины за 1963 год. Модель получила название модель Ходжкина-Хаксли
В 2005 году Томасом Геймбургом и Анрю Д. Джексоном предложена солитонном модель, основанная на предположении, что сигнал по нейронам распространяется в виде солитонов — устойчивых волн, распространяющихся по клеточной мембране.
Влияние некоторых веществ на потенциал действия
Некоторые вещества органического или синтетического происхождения могут блокировать образование или прохождения ПД:
- Батрахотоксин найден у некоторых представителей рода листолазов. Устойчиво и необратимо повышает проницаемость мембран для ионов натрия.
- Понератоксин был найден в муравьях рода Paraponera. Блокирует натриевые каналы.
- Тетродотоксин найден в тканях рыб семейства Скелезубови, из которых готовят японский деликатес Фугу. Блокирует натриевые каналы.
- Механизм действия большинства анестетиков (Прокаин, Лидокаин) базируется на блокировании натриевых каналов и соответственно на блокировании проведении импульсов по чувствительным нервным волокнам.
- 4-Аминопиридин — обратно блокирует калиевые каналы, удлиняет срок потенциала действия. Может использоваться в терапии рассеянного склероза.
- ADWX 1 — обратно блокирует калиевые каналы. В условиях опыта облегчал течение острого рассеянного энцефаломиелита у крыс.
Изображения по теме
| Рис. 19-26. Измерение тока через открытый канал ацетилхолинового рецептора при разных значениях мембранного потенциала. С помощью таких измерений можно установить ионную селективность каналов. Ток, переносимый через открытый канал ионами определенного вида, будет изменяться при изменении мембранного потенциала определенным образом в зависимости от вида иона и градиента его концентрации по обе стороны мембраны. Зная градиенты концентраций основных присутствующих ионов, можно определить ионную селективность канала путем простого измерения зависимости ток/напряжение более полную информацию можно получить в результате повторных измерений при других концентрациях иона. А. Зарегистрированный с помощью метода пэтч-клампа ток, проходящий через одиночный канал, находящийся в растворе с фиксированной концентрацией ацетилхолина, при трех различных значениях мембранного потенциала. В каждом случае канал случайным образом переходит из закрытого состояния в открытое и обратно, но при некотором значении мембранного потенциала, которое называют потенциалом реверсии, гок равен нулю даже тогда, когда канал открыт. В данном случае потенциал реверсии близок к О мВ. Б. Такое же явление можно наблюдать, измеряя после одиночной стимуляции нерва общий ток через больщое количество одиночных каналов с ацетилхолиновым рецептором, находящихся в постсинаптической мембране нервно-мыщечного соединения. На графиках показаны изменения этого гока, измеренного с помощью внутриклеточных электродов в условиях фиксации напряжения. Каналы открываются при коротком воздействии ацетилхолина, но если мембранный потенциал поддерживается на уровне потенциала реверсии, го ток равен нулю. Поскольку открытые каналы проницаемы как для Na . так и для К . а значения электрохимических движущих сил для этих ионов различны, нулевой ток в действительности соответствует уравновещенным и направленным навстречу друг другу токам Na и К . (Эти каналы проницаемы и для Са , но ток, переносимый ионами кальция, очень мал, так как их концентрация низка.) |
Объяснение механизма обратного взимания НДС — Определение, пример и др.
На практике обратное начисление упрощает уплату НДС (по крайней мере, со стороны поставщика) и снижает соответствующие бюрократические расходы . Европейский Союз создал эту налоговую концепцию для борьбы с такими нарушениями, как налоговое мошенничество в рамках единого европейского рынка. С момента его внедрения в реформу системы налога на добавленную стоимость ЕС в 1993 году он особенно успешно боролся с так называемыми «схемами пропавших без вести торговцев», когда не облагаемые налогом трансграничные перевозки используются для уклонения от уплаты НДС.По оценкам, этот метод теряет несколько миллиардов евро в год.
Пример : Товары импортируются из Соединенных Штатов во Францию, где предприниматель A приобретает и впоследствии передает их предпринимателю B в Великобритании. Предприниматель А выставляет счет без НДС. Предприниматель B продает товары от предпринимателя A предпринимателю C в Великобритании. Однако на этот раз предприниматель Б выставляет счет с соответствующими цифрами НДС. НДС возвращается предпринимателю C налоговой службой в виде предналога.Однако вместо того, чтобы платить НДС в налоговую инспекцию, предприниматель Б исчезает с рынка до того, как наступит срок уплаты налога. С этого момента его называют «пропавшим трейдером». Затем предприниматель C продает товары обратно предпринимателю A, и весь процесс повторяется.
Несмотря на то, что поставщики не производят уплату НДС в случае мошенничества со стороны торговцев, они, тем не менее, требуются бенефициарами в виде предварительных налогов. Механизм обратного начисления предотвращает такие ситуации, сохраняя все НДС и входящие налоги под одной крышей.Если получатель хочет получить возмещение в виде предналога от налоговой инспекции, он также должен четко указать НДС, включенный в счет. Поскольку обратные сборы делают налоговые системы менее уязвимыми для мошенничества , многие европейские отрасли в настоящее время рассматривают эту процедуру как обязательную часть любой транзакции. В прошлом строительная отрасль и поставщики планшетов и мобильных телефонов были очень частыми целями таких мошеннических операций.
.
обратный заряд — это … Что такое обратный заряд?
reverse-charge — UK US прилагательное [перед существительным] UK (сбор в США) ► КОММУНИКАЦИИ, используемые для описания телефонного звонка, который оплачивается лицом, которое его получает: »Мы сделали обратный звонок в Новую Офис в Йорке… Финансовые и коммерческие условия
Обратный платеж — может означать: * При размещении телефонных звонков обратный платеж также известен как звонок по сбору. * В случае перезаряжаемых батарей ошибка при использовании или зарядке может привести к обратной зарядке… Wikipedia
обратное начисление — В контексте налога на добавленную стоимость (НДС), когда определенные услуги со стандартным тарифом (перечисленные в Приложении 5 Закона о налоге на добавленную стоимость 1994 г.) предоставляются лицу в Великобритании торговцем за пределами Великобритании (внутри или за пределами ЕС), получатель из Великобритании…… Юридический словарь
Обратный заряд — Das Abzugsverfahren (int.: Обратный заряд) ist eine Sondernorm im Umsatzsteuerrecht, die den Übergang der Steuerschuldnerschaft regelt. Nach derzeit geltendem Umsatzsteuergesetz muss ein Leistungserbringer die Umsatzsteuer vom Leistungsempfänger…… Deutsch Wikipedia
обратный заряд — прил. Брит: ↑ сбор, 2 обратный заряд телефонный звонок… Полезный английский словарь
обратный платеж — Обычно поставщик товаров или услуг должен отчитываться перед Налоговой и таможенной службой Великобритании по уплате НДС, причитающегося с поставок.Однако в определенных ситуациях НДС должен учитывать покупатель. Также известен как налоговый сдвиг. HM Customs &… Финансовые и коммерческие условия
обратная оплата — / rəˈvɜs tʃadʒ / (скажем, ruh vers chahj) прилагательное (телефонного звонка), оплаченное лицом, принимающим звонок… Словарь австралийского английского
звонок с обратной оплатой — Великобритания США существительное [исчисляемое] [звонок с обратной оплатой в единственном числе звонки с обратной оплатой во множественном числе] британский телефонный звонок, за который человек, которому вы звоните, соглашается оплатить.Американское слово — колл-колл. Тезаурус: телефонные звонки и типы телефонов…… Полезный английский словарь
вызов с обратным начислением — вызовы с обратным начислением N COUNT Звонок с обратным начислением — это телефонный разговор, который оплачивается лицом, которое получает вызов, а не лицом, которое звонит. [BRIT] (в AM, используйте сборный звонок)… Английский словарь
обратный звонок — британское / американское существительное [исчисляемое] Формы слова обратный звонок: единственное число обратный звонок множественный обратный звонок британский телефонный звонок, за который человек, которому вы звоните, соглашается заплатить.Американское слово — это звонок… Английский словарь
обратный вызов — обратный, стихотворный вызов счет существительное BRITISH a COLLECT CALL… Использование слов и фраз в современном английском языке
.
Возврат НДС — Финансы | Динамика 365
- 5 минут на чтение
В этой статье
Важно
Dynamics 365 for Finance and Operations превратилась в специализированные приложения, которые помогут вам управлять конкретными бизнес-функциями.Дополнительные сведения об этих изменениях см. В Руководстве по лицензированию Dynamics 365.
В этом разделе описывается общий подход к настройке обратного налога на добавленную стоимость (НДС) для Саудовской Аравии, Сингапура и европейских стран.
Reverse Charge — это налоговая схема, которая перекладывает ответственность за учет и отчетность по НДС с продавца на покупателя товаров и / или услуг. Таким образом, получатели товаров и / или услуг сообщают как выходной НДС (в роли продавца), так и входящий НДС (в роли покупателя) в своем отчете по НДС.
В некоторых странах или регионах схема обратного начисления применяется только для некоторых товаров и / или услуг, и существуют дополнительные условия или пороговые значения для сумм продаж. В других странах или регионах ответственность за уплату НДС зависит от статуса поставщика и покупателя. Если покупатель обязан уплатить НДС, этот факт должен быть четко указан в счете, выставляемом поставщиком. Например, счет-фактура должен включать слова «Обратный платеж» и указывать, какие позиции относятся к схеме обратного платежа.
Чтобы применить обратный заряд, необходимо выполнить следующую настройку.
Настройка налоговых кодов
Мы рекомендуем использовать отдельные налоговые коды для операций продаж и операций покупки.
| Налоговый код для продаж | Создайте налоговый код для операций по продажам с обратным начислением ( Налог > Косвенные налоги > Налог с продаж > Налоговые коды ). |
| Налоговый код для покупок | Создайте положительные и отрицательные коды налога для обратного начисления НДС для покупок ( Налог > Косвенные налоги > Налог с продаж > Коды налога с продаж ).
Для получения дополнительной информации см. Следующий раздел «Настройка налоговых групп и налоговых групп товаров.« |
Настройка налоговых групп и налоговых групп товаров
Мы рекомендуем использовать отдельные налоговые группы для операций продаж и операций покупки.
| Налоговые группы для продаж | Создайте налоговую группу для операций продаж с обратным начислением ( Налог > Косвенные налоги > Налог с продаж > Налоговые группы ). На вкладке Настройка включите налоговый код для обратного начисления в эту группу.Установите флажки Exempt и Reverse charge для налогового кода. |
| Налоговые группы для покупок | Создайте налоговую группу для операций покупки с обратным начислением ( Налог > Косвенные налоги > Налог с продаж > Налоговые группы ). На вкладке Настройка включите в эту группу как положительные, так и отрицательные коды налога с продаж. Установите флажок Обратный сбор для кода налога с продаж с отрицательным значением. |
| Позиция налоговых групп | Создайте или обновите налоговую группу номенклатур с кодом налога с отрицательным значением ( Налог > Косвенные налоги > Налог с продаж > Налоговые группы номенклатуры ). Вы должны назначить налоговую группу номенклатур по умолчанию для продуктов и категорий, которые подлежат обратному начислению. |
Настройка групп обратного начисления
На странице Группы позиций обратных начислений ( Налог > Настройка > Налог с продаж > Группы позиций обратных начислений ) вы можете определить группы продуктов или услуг или отдельные продукты или услуги, которые реверсируются заряд может быть применен к.Для каждой группы товаров обратного начисления определите список товаров, групп товаров и категорий для продаж и / или покупок.
Настройка правил обратного начисления
На странице Правила обратного начисления ( Налог > Настройка > Налог с продаж > Правила обратного начисления ) вы можете определить правила применимости для целей покупки и продажи. Вы можете настроить набор правил применимости обратного начисления. Для каждого правила установите следующие поля:
- Тип документа — Выберите Заказ на поставку , Журнал накладных поставщика , Заказ на продажу , Счет-фактура с произвольным текстом , Журнал накладных клиента и / или Счет-фактура поставщика .
- Страна / регион, тип партнера — выберите Внутренний , ЕС или Иностранный . В качестве альтернативы, если правило может применяться ко всем торговым партнерам, независимо от страны или региона их адреса, выберите Все .
- Адрес доставки внутри страны — установите этот флажок, чтобы применить правило к доставке в пределах одной страны или региона. Этот флажок нельзя установить для типов документов Журнал накладных поставщика и Журнал накладных клиентов .
- Группа позиций обратных начислений — Выберите группу, к которой можно применить правило.
- Пороговая сумма — Схема обратного начисления применяется к счету только в том случае, если стоимость товаров и / или услуг, включенных в группу товаров обратных начислений, превышает предел, указанный здесь.
Вы также можете использовать поля Дата вступления в силу и Срок действия , чтобы определить период действия правила.
Кроме того, вы можете указать, будет ли отображаться уведомление и обновляться ли строка документа с использованием группы налога обратного начисления по умолчанию, если выполняется условие для этой строки документа.Доступны следующие варианты:
- Нет — строка документа не обновляется.
- Подсказка — появляется уведомление, подтверждающее, что обратная оплата может быть применена.
- Установить — строка документа обновляется без дополнительного уведомления.
Установить параметры по умолчанию
Чтобы включить функцию обратного начисления НДС, на странице Параметры главной книги на вкладке Обратное начисление установите для параметра Включить обратное начисление значение Да .В полях Налоговая группа заказов на покупку и Налоговая группа заказов на продажу выберите налоговые группы по умолчанию. Когда выполняется условие применимости обратных начислений, строка заказа на продажу или покупку обновляется этими налоговыми группами.
Обратное начисление по счету-фактуре продажи
Для продаж по схеме обратного сбора продавец не взимает НДС. Вместо этого в счете-фактуре указываются как позиции, которые подлежат обратному начислению НДС, так и общая сумма обратного НДС.
При разноске счета-фактуры с обратным начислением налоговые проводки имеют направление налога к уплате и нулевой налог с продаж, а также установлен флажок Обратный сбор .
Обратный платеж по счету-фактуре покупки
Для покупок по схеме «Обратный сбор» покупатель, получивший счет-фактуру с обратным начислением, действует как покупатель и продавец для целей учета НДС.
При разноске накладной покупки с обратным начислением создаются две налоговые проводки.Одна транзакция имеет направление Налог к получению . Другая транзакция имеет направление Налог к уплате , и установлен флажок Обратный сбор .
На следующем снимке экрана одна транзакция имеет направление Налог к получению , а другая транзакция имеет направление Налог к оплате .
.
Механизм обратного платежа под НДС
Что такое обратный заряд?
В обычном бизнесе поставщик поставляет товары и взимает НДС от имени клиентов, который позже уплачивается государству. Согласно механизму обратной оплаты, покупатель или конечный покупатель платит налог напрямую государству.
Поставщик не обязан платить НДС по импортируемым товарам, поэтому обязанность сообщать о сделке по НДС перекладывается с продавца на получателя.Получатель должен будет регистрировать НДС на покупки (входящий НДС) и НДС на продажи (исходящий НДС) в своей декларации по НДС каждый квартал.
Зачем нужна обратная зарядка?
Если поставщик не ведет бизнес в ОАЭ, отслеживание его деловых операций и обеспечение соблюдения НДС становится утомительным для FTA. Следовательно, получатели, являющиеся резидентами ОАЭ, вынуждены платить НДС на основе обратного начисления. Механизм обратной оплаты в основном используется для трансграничных транзакций.Это освобождает поставщиков-нерезидентов от бремени регистрации и учета НДС по месту нахождения их покупателей.
Когда применяется обратный сбор?
Обратный платеж применяется в следующих случаях:
- Импорт товаров / услуг из других стран ССЗ и не входящих в ССЗ. Поставщик этих товаров / услуг должен находиться в другой стране, и он может иметь или не вести бизнес в ОАЭ.
- Покупка товаров из обозначенной зоны
- Поставка золота и алмазов
- Покупка золота и алмазов для перепродажи или дальнейшего производства / производства
- Поставка углеводородов для перепродажи зарегистрированным поставщиком зарегистрированному получателю в ОАЭ
- Поставка сырой / рафинированной нефти зарегистрированным поставщиком зарегистрированному получателю в ОАЭ
- Поставка переработанного / необработанного природного газа зарегистрированным поставщиком зарегистрированному получателю в ОАЭ
- Производство и распределение любой формы энергии, поставляемой зарегистрированным поставщиком зарегистрированному получателю в ОАЭ
Примечание. Механизм обратной оплаты не применяется к экспорту золота и алмазов, поставкам инвестиционных драгоценных металлов (платина или золото с чистотой более 99%, которые торгуются на мировых рынках) и экспорту продуктов, основным компонентом которых является золото или бриллианты.
Как работает механизм обратной зарядки?
Чтобы лучше понять механизм, рассмотрим пример использования импортированной службы:
Г-н Абдул зарегистрирован в качестве плательщика НДС в ОАЭ, но пользуется услугами бухгалтера по имени Сэм, который находится в Италии. Поскольку Сэм не зарегистрирован в ОАЭ, ему не нужно подавать какие-либо декларации в ОАЭ или платить налоги ОАЭ. Поскольку г-н Абдул приобрел услуги у поставщика, не находящегося в ОАЭ, он должен будет указать обратный платеж в своей соответствующей декларации по НДС.В этом примере, поскольку получатель учитывает НДС в соответствии с механизмом обратного начисления, местом поставки для целей НДС является ОАЭ.
Что мне делать, если я являюсь получателем, участвующим в транзакции обратного списания?
Вы должны рассчитать сумму налога, подлежащего уплате государству, самостоятельно учесть сумму НДС как исходящий налог во время покупки и затем указать ее в своей декларации по НДС. Если возможно, вы можете потребовать входной кредит. Убедитесь, что у вас есть необходимые документы, такие как счета, для использования в будущем.
Какие требования к механизму обратного заряда?
- Получатель товаров или услуг должен быть зарегистрирован в качестве плательщика НДС.
- Каждый зарегистрированный владелец бизнеса должен вести надлежащий учет своих запасов, за которые взимается обратная плата.
- В счетах-фактурах, квитанциях и квитанциях на возврат следует указывать, взимается ли налог, подлежащий уплате по данной конкретной транзакции, путем обратного взимания.
.